Plan du chapitre
- Introduction et problématique
- I – La conservation des génomes au sein d’un individu : stabilité génétique et évolution clonale
- II – Le brassage des génomes au cours de la fécondation et de la méiose
- III – Les accidents génétiques de la méiose
- Conclusion
- Exercices
- Entrainement sur des exercices de type bac
- Pour aller plus loin

Cours, schémas et fiche mémo en version imprimable
Lien vers le padlet des arguments
lien à venir (n’hésitez pas à le signaler à vos profs si le chapitre est commencé et ce lien est encore absent)
Activités du chapitre
– Activité 0 : Les cellules sanguines, une mosaïque de clones cellulaires + Correction
– Activité 1 : Les étapes de la méiose (révisions)+ Correction
– Activité 2 : les expériences de Mendel + Activité 2 – corrigé
– Activité 3 – Des croisements en botanique (étude des tableaux de croisements) + correction
– Activité 3BIS – les tableaux de croisements dans l’exemple des chevaux Alezan + correction
– Activité 4 – le test-cross (TD) + le diaporama de cette séance
– Activité 5 : Les accidents de la méiose
– Activité 6 : La mucoviscidose
– Activité 6 bis : un exemple de famille multigénique : les globines + Activité 6 bis – corrigé (non fait en 2025)
– Activité 7 : Une protéine « poison » du grain de pollen + corrigé (barème/schéma)
Ressources du TP 5 – Flyroom
Ressources du TP6 – Cafouillage au laboratoire
– fiche technique du logiciel MESURIM
– fiche du TP6 – Cafouillage au laboratoire
– corrigé du TP6 (Merci à Flore et Fatima)
– corrigé du TP6 (version 2)
Ressource du TP 7 – Variations génétiques et phénotypiques
Le lien vers le logiciel Geniegen 2 en ligne
Corrigé du TP (merci aux 4 élèves anonymes de l’année 2022-2023)
La vidéo présentant la technique ELISA
Ressources du TP6bis – Mucoviscidose et conseil génétique (non réalisé en 2025)
– fiche du TP6A – Myopathie de Duchenne
– fiche du TP6B Mucoviscidose et conseil génétique
– lien vers GENIEGEN2
Fichiers séquence à utiliser :
– famille 1
– famille 2
– famille 3
– famille MUCO
Table du code génétique :

Exercices du chapitre

Ce module d’exercices a été conçu à partir du cours du prof de cette page à l’aide d’une intelligence artificielle (Nolej IA)
Ce site a été réalisé par Mme et Mr ESTHER

Introduction

Par reproduction sexuée, deux parents donnent naissance à de nouveaux individus dont le génome est unique. Dans ce chapitre, nous allons mettre en évidence les mécanismes de la méiose (production des gamètes et de la fécondation (union des gamètes) qui expliquent l’origine des génotypes de chaque individu.
Problématique
Comment les divisions cellulaires (mitose et méiose) et les mécanismes de la fécondation aboutissent-ils à la formation de nouveaux génotypes propres à chaque individu ?
ou
Quels sont les processus qui expliquent la diversité génotypique des individus ?
Ce site a été réalisé par Mme et Mr ESTHE

I – La conservation des génomes au sein d’un individu : stabilité génétique et évolution clonale
Une cellule qui subit une succession de mitoses donne naissance à un ensemble de cellules toutes génétiquement semblables appelées clones. Ces clones forment des ensembles de cellules séparées (exemple des cellules sanguines) ou des ensembles de cellules associées formant des tissus (exemple des cellules de la peau).
Du fait de l’absence d’échanges génétiques avec l’extérieur, la diversité génétique dans ces lignées clonales résulte d’accidents génétiques tels que les mutations. Ces modifications (si elles ne sont pas réparées) sont transmises à l’ensemble des cellules dérivant du mutant et peuvent devenir héréditaires si elles surviennent dans une cellule de la lignée germinale.
Rappel : les mutations peuvent soit être sans effet, soit avoir un effet négatif, soit être à l’origine de caractères nouveaux susceptibles d’être sélectionnés au cours de l’évolution.
On peut donc considérer un individu comme une mosaïque de clones avec des génomes qui diffèrent très légèrement d’une lignée à l’autre du fait des mutations.
Voici un exemple de représentation des mosaïques de clones (Source : https://science.sciencemag.org/content/354/6312/557/tab-figures-data)
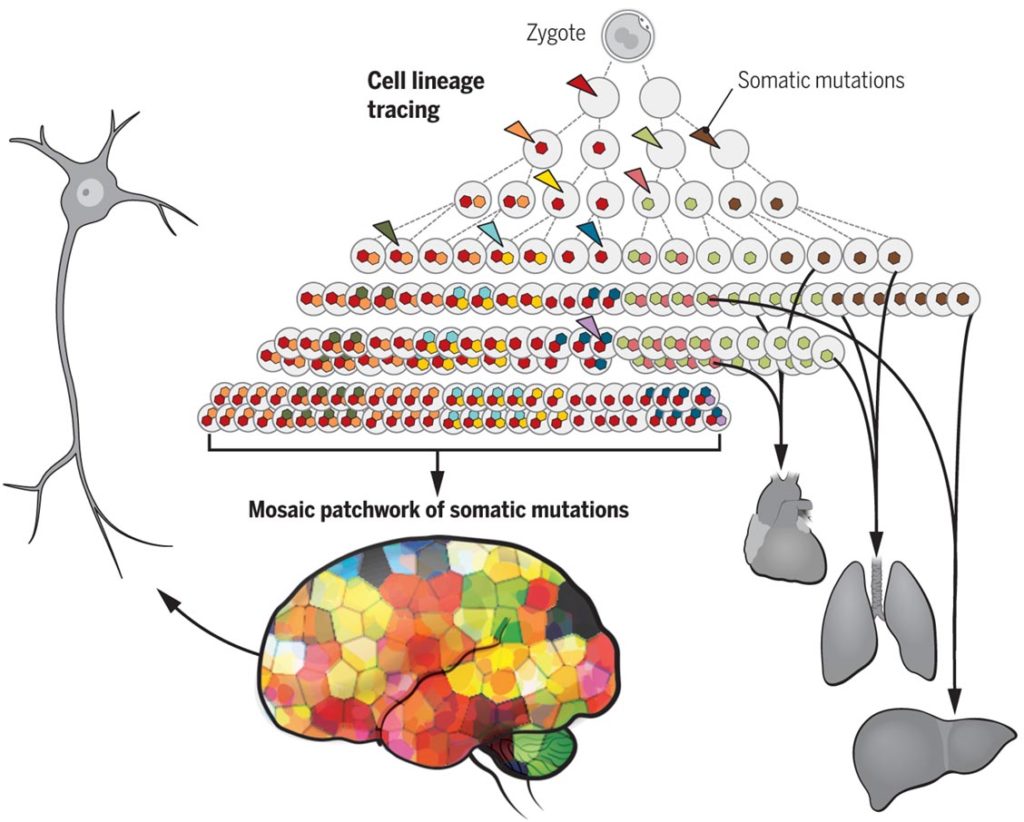
Voici un autre exemple de représentation des mosaïques de clones (Source : Science)

Pour aller plus loin – Une étude de « Science » publiée en juin 2019
Ce site a été réalisé par Mme et Mr ESTHER
II – Le brassage des génomes au cours de la fécondation et de la méiose
1 – La méiose : la formation de gamètes présentant une grande diversité génétique (à partir d’un génome unique)
|
Rappels sur la méiose
La méiose est toujours précédée d’une phase de réplication de l’ADN. La méiose est un ensemble de 2 divisions cellulaires successives et inséparables :
|


Exemple de schéma possible d’une méiose possible d’une cellule 2n = 4 avec 3 gènes.
La méiose fait passer d’une phase diploïde (cellules à 2n chromosomes) à une phase haploïde (cellule à n chromosomes). Une cellule-mère (2n) au départ donne 4 cellules filles (n). Lors de la méiose, le programme génétique n’est pas conservé : les cellules-filles n’ont pas le même matériel génétique que leur mère.
Deux types de brassages existent naturellement au cours de la méiose :
- le brassage interchromosomique : il correspond aux différentes combinaisons de chromosomes reçus par les cellules filles du fait de la migration aléatoire (au hasard) des chromosomes de chaque paire lors de la 1ère division de méiose. Chaque chromosome d’une même paire n’ayant pas les mêmes allèles pour chaque gène (30% de polyallélisme dans l’espèce humaine).

- le brassage intrachromosomique : il correspond à des échanges de fragments d’ADN entre les chromatides d’une même paire de chromosomes, par crossing-over ou enjambement. Les chromosomes ainsi formés sont dits recombinés.

Schéma montrant les conséquences d’un crossing-over sur la répartition des allèles entre les chromatides de chromosomes homologues

Ces deux types de brassages se produisent à chaque méiose dans l’espèce humaine. Ainsi, il y a un grand nombre de combinaisons génétiques possibles dans les gamètes ; d’autant plus grand que le nombre de gènes présents à l’état hétérozygote est grand chez les parents.
2 – La fécondation : rencontre au hasard et fusion de deux gamètes porteurs d’un lot d’allèles
Le processus de fécondation est universel : le noyau haploïde d’un gamète mâle s’unit au noyau haploïde d’un gamète femelle. La fusion des deux gamètes aboutit à un zygote. Cette fusion des 2 noyaux est appelée caryogamie. Cela forme une cellule-œuf. Les paires de chromosomes homologues sont ainsi reconstituées (dans chaque paire, un chromosome est issu du père, l’autre de la mère). Chaque zygote contient ainsi une combinaison unique et nouvelle d’allèles.
|
Illustrations – Piste possible pour un argument (mathématique !)
|
La diversité génétique potentielle des zygotes est donc immense. Cependant, seule une partie de ces zygotes est viable et se développe.
| Illustrations : Les arrêts spontanés de grossesse dont font partie les fausses-couches s’expliquent en partie par l’arrêt de développement de certains zygotes/embryons non viables du fait d’anomalies génétiques. D’après Santé Publique France, 50% des avortements spontanés au premier trimestre sont liés à des anomalies chromosomiques. |
Une vidéo bonus sur la méiose
Des précisions sur Gregor Mendel et les lois de l’hérédité
Ce site a été réalisé par Mme et Mr ESTHER
III – Les accidents génétiques de la méiose
Des anomalies peuvent survenir au cours de la méiose : crossing-over inégal, anomalies de la séparation du matériel génétique au cours des divisions de méiose… Ces accidents sont souvent létaux (non viable pour le zygote formé) mais ils engendrent parfois une diversification importante des génomes et jouent un rôle essentiel dans l’évolution biologique (familles multigéniques).
1 – Les anomalies lors de la séparation du matériel génétique
La méiose est une étape clé de la reproduction sexuée, particulièrement chez les espèces à dominante diploïde où elle assure la production des gamètes. Si un gamète est anormal, la cellule-œuf résultant de la fécondation peut avoir un nombre aberrant de chromosomes. C’est le cas de la trisomie (chromosome en 3 exemplaires) ou de la monosomie (chromosome en 1 exemplaire). Ces maladies sont souvent graves ce qui montre la nécessité du maintien du caryotype.
Ces anomalies peuvent survenir au cours de la 1 ère division de méiose – il y a non-séparation des chromosomes homologues – ou au cours de la 2 ème division – il y a non-disjonction des chromatides sœurs.

Schéma montrant les 2 anomalies possibles de méiose pouvant conduire à des trisomies ou des monosomies (après fécondation avec un autre gamète normal) – Exemple de la trisomie 21
|
Exemples : Les trisomies 21 sont les anomalies de séparation du matériel génétique les plus connues. |
2 – Les anomalies lors des crossing-over
Des anomalies peuvent aussi survenir au cours des crossing-over. Si les portions échangées sont inégales, un crossing-over peut parfois aboutir à une duplication d’un gène. La plupart du temps, ces mécanismes sont sources de troubles mais ils sont parfois sources de diversification du vivant, par exemple à l’origine des familles multigéniques.
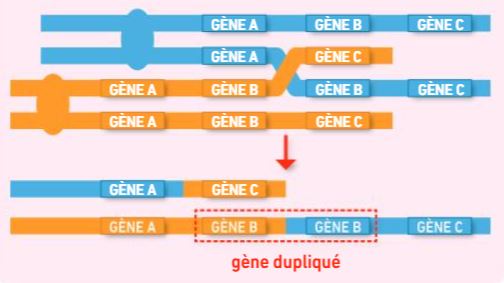
Schéma d’un crossing-over inégal entrainant pour une chromatide, une perte de gène, pour l’autre un gène dupliqué (en 2 copies) – Source : Bordas
Au sein d’une même espèce, on remarque des similitudes entre plusieurs gènes (ex : myoglobine, hémoglobine). Tous ces gènes présentant une forte ressemblance forment une famille multigénique.
L’existence de ces similitudes est interprétée comme le résultat d’une (ou plusieurs) duplication(s) d’un gène ancestral du fait de crossing-over inégaux.
La divergence entre les gènes d’une même famille multigénique s’explique par l’accumulation de mutations (différentes selon les duplicats). Dans certains cas, ces processus peuvent conduire à l’acquisition de gènes correspondant à de nouvelles fonctions.

Schéma les conséquences des duplications de gènes liés à des crossing-over inégaux sur l’évolution des espèces – Source : Bordas
|
Exemples : |
Conclusion
Version 1 – Les mécanismes de la méiose et de la fécondation sont sources de très grande diversité dans la production et la rencontre des gamètes. Ils aboutissent ainsi à des génotypes uniques à chaque individu. De plus, les événements de mutations et de mitose conduisent à une modification de génotype des différentes cellules constituant un individu au cours de sa vie. La reproduction sexuée génère donc une très grande diversité parmi les individus d’une même espèce.
Version 2 – Lors de la reproduction sexuée se mettent en place des brassages qui sont sources d’une diversité génétique quasi infinie au sein d’une même espèce. Cela explique la diversité génétique à l’échelle des individus dans une population.
Par ailleurs, au sein d’un même individu, on constate également une certaine diversité génétique à l’échelle cellulaire. Elle est due aux mutations somatiques qui se produisent au cours de la vie.
Question : quels sont les autres mécanismes qui permettent de créer de la diversité génétique au sein d’une espèce ?
Ce site a été réalisé par Mme et Mr ESTHER
Ce site a été réalisé par Mme et Mr ESTHER

Quelques QCM (proposés par un autre site) pour vous entraîner. Attention il déclenche des points négatifs (-0.5) en cas d’erreurs !
QCM1 – Je vérifie si je sais identifier les gènes, les allèles, la notion hétérozygote, homozygote et si je sais étudier les croisements.
QCM2 – Je vérifie si je sais faire le lien et la différence entre génotype et phénotype.
QCM3 – Je vérifie si je maîtrise les termes haploïdes et diploïdes.
Ce site a été réalisé par Mme et Mr ESTHER
Voici quelques exemples possibles de sujets de bac sur ce chapitre :
- exercices de type 1 (proposés en AP SVT)
- exercice de type 2
- exercices de type ECE1 (proposés en AP SVT)

Un podcast sur un travail de recherche sur la trisomie 21 de Marthe Gautier