Plan du chapitre
- Introduction et problématique
- I – L’évolution des espèces et le modèle de l’équilibre d’Hardy-Weinberg
- II – La notion d’espèce, la spéciation et les techniques modernes de séquençage génétique
- Exercices
- Entrainement sur des exercices de type bac
- Pour aller plus loin

Lien vers le padlet des arguments
Activités du chapitre
TP – La génétique de la tolérance au lactose dans les populations humaines
TP – Evolution de la coloration du pelage chez les souris des plages
- Fiche TP – Evolution de la coloration du pelage chez les souris des plages
- Tableur Calc des données des documents
- Modèle Edu’modèles 1 : Souris-jaune_vs_Souris-grise_LAVE.modele
- Modèle Edu’modèles 1 : Souris-jaune_vs_Souris-grise_SABLE.modele
- Lien vers le logiciel Edu’modèles
Fiche de travail pour le Poster de la partie A
TP – Evolution de populations d’éléphants (sélection naturelle et dérive génétique)
- Fiche du TP
- Les modèles à ouvrir dans Edu’modèles
- Modèle Edu’modèles 1 : Elephants – HW.modele
- Modèle Edu’modèles 2 : ElephantsAddo.modele
- Modèle Edu’modèles 3 : ElephantsLuangwa.modele
Les éléments de correction des modèles :
- Modèle Edu’modèles 2 : ElephantsAddo – corrigé.modele
- Modèle Edu’modèles 1 : ElephantsLuangwa – corrigé.modele

Ce module d’exercices a été réalisé à partir d’une IA (Nolej IA) et des cours de Mr et Mme Esther. Des erreurs et/ou des maladresses de formulation peuvent persister.
Ce site a été réalisé par Mme et Mr ESTHER

Introduction
Au sein d’une espèce, dans les différentes populations, les allèles de chaque gène sont transmis de génération en génération selon les processus de méiose et fécondation. Cependant, dans les populations naturelles, inexorablement, des différences génétiques apparaissent d’une population à l’autre. Ces modifications peuvent parfois aboutir à la formation de nouvelles espèces.
Vidéo de la partie pierre-feuille-ciseaux des lézards !
https://www.facebook.com/watch/?v=2146296308915944
Problématique
Quels sont les mécanismes de l’évolution des génomes au sein des populations ?
Ce site a été réalisé par Mme et Mr ESTHER
I – L’évolution des espèces et le modèle de l’équilibre d’Hardy-Weinberg
1) Un équilibre théorique mis en évidence par Hardy et Weinberg
Le modèle mathématique de Hardy-Weinberg a été énoncé au début du 20ème siècle indépendamment par le mathématicien Godfrey H. HARDY et par le médecin gynécologue Wilhelm WEINBERG.
Le modèle de Hardy-Weinberg postule que, pour une population qui respecte quelques conditions* :
- les fréquences alléliques sont stables d’une génération à l’autre ;
- les fréquences alléliques et les fréquences génotypiques sont liées par une équation mathématique simple :
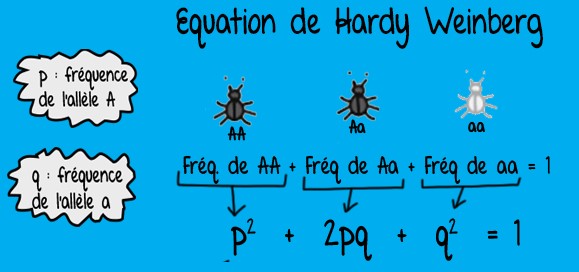
*Ce modèle s’applique dans des conditions particulières :
- organisme diploïde (deux allèles pour chaque gène) se reproduisant par reproduction sexuée
- taille de la population infinie
- rencontre au hasard des partenaires et des gamètes pour la reproduction (panmixie)
- population fermée (sans mouvement migratoire)
- aucune force évolutive (mutation, dérive, sélection, migration)
Le modèle de Hardy-Weinberg décrit une cas théorique d’une population qui n’évolue pas ! On parle d’équilibre de Hardy-Weinberg.
Il est possible d’élaborer des modèles numériques (type Edu-modèle ou Evolution des fréquences alléliques) qui s’appuient sur les règles de séparation des allèles lors de la méiose et de leur réassociation au hasard lors de la fécondation et qui permettent de calculer les effectifs des génotypes de la descendance.
En appliquant les règles de Hardy-Weinberg, il est possible de retrouver cette constance des fréquences alléliques prédite par le modèle théorique de Hardy et Weinberg.
Cependant, dans les populations naturelles, les conditions de l’équilibre de HW sont rarement vérifiées. Les modèles peuvent ainsi être adaptés afin de représenter les évolutions observées dans la population naturelle.
2) Les écarts à l’équilibre ou à la structure de Hardy-Weinberg
Au sein des populations, différentes forces évolutives agissent et font varier les fréquences alléliques et génotypiques :
- Des mutations apparaissent spontanément chez les individus ce qui entrainent la formation de nouveaux allèles et modifie ainsi la fréquence des allèles préexistants
- Exemple de mutations (celles de la mucoviscidose – gène CFTR)
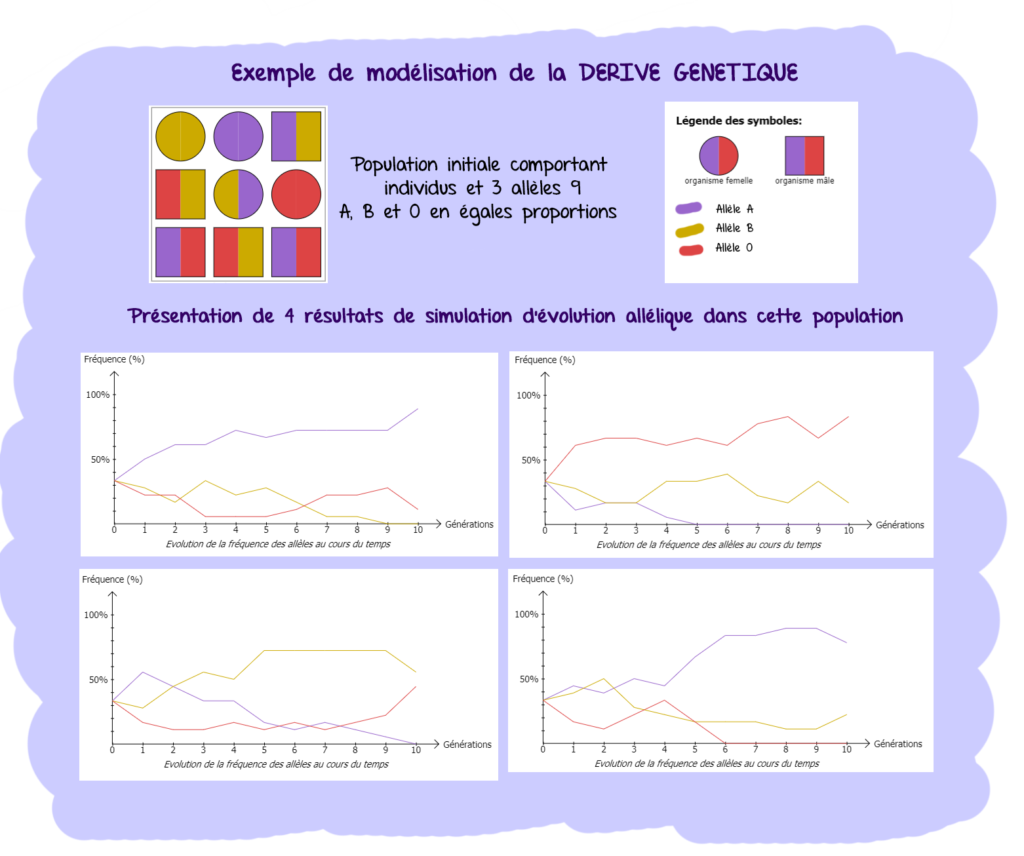
- Dans la nature, les populations ne sont pas de taille infinie et la fréquence des allèles de population de taille limitée varie sous l’effet du hasard : c’est la dérive génétique
- Exemple de la prédominance de gènes de prédisposition à l’obésité dans les populations micronésiennes
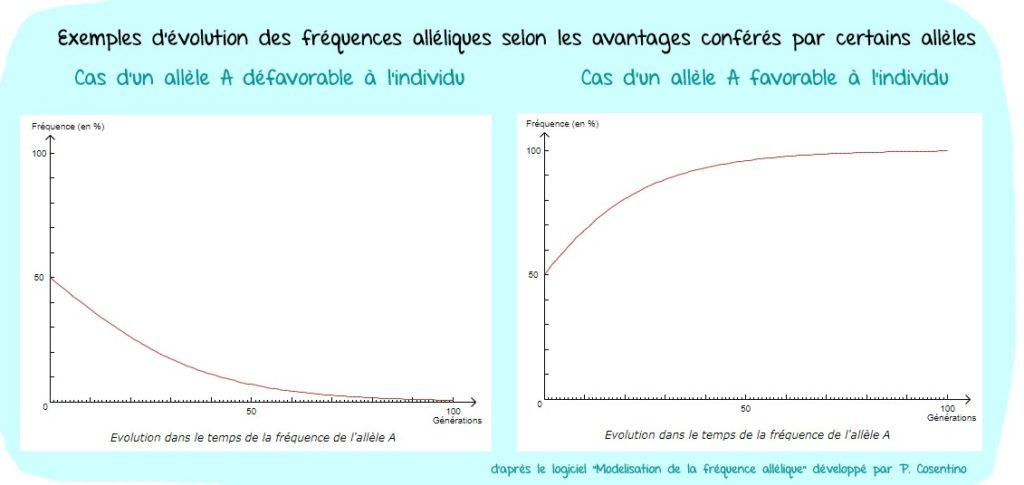
- Certains allèles confèrent un avantage (ou un désavantage) aux individus le possédant, soit pour leur survie, soit pour leur reproduction : c’est la sélection naturelle. Certains allèles se répandant davantage dans la population alors que d’autres ont tendance à disparaitre.
- Exemple des défenses des éléphants (TP 7)
- L’existence de préférence sexuelle entre les partenaires influence aussi la fréquence des allèles au cours du temps
- Exemple des défenses des éléphants (TP 7)
- Les migrations de populations souvent de taille réduite influencent fortement la fréquence des allèles au cours du temps.
- Exemple de la porphyrie variegata (maladie affectant de nombreux Afrikaners), exemples des fréquences des allèles du groupe sanguin selon les populations humaines

Ce site a été réalisé par Mme et Mr ESTHER
II – La notion d’espèce, la spéciation et les techniques modernes de séquençage génétique
Rappels : la notion d’espèce
La définition ou le concept le plus reconnu d’une espèce a été proposé par Ernst MAYR* en 1942: « une espèce est un groupe de populations naturelles au sein duquel les individus peuvent, réellement ou potentiellement, échanger du matériel génétique ; toute espèce est séparée des autres par des mécanismes d’isolement reproductif ». Ce concept est associé à la nécessité sur le terrain de reconnaître/nommer les êtres vivants observés. D’un point de vue opérationnel, pour distinguer deux espèces ont s’appuie sur deux critères : les ressemblances morpho-anatomiques et l’interfécondité. Cela fonctionne pour un grand nombre d’espèces mais l’interfécondité est souvent difficile à établir (ex : pour les fossiles) et les ressemblances morpho-anatomiques ne traduisent pas toujours les différences/ressemblances génétiques entre populations (ex : le canard colvert**).
Comme on l’a montré avec le modèle de Hardy-Weinberg, les populations des différentes espèces sont en constante évolution sous l’effet combiné des forces évolutives (sélection naturelle/sexuelle, dérive, etc) et des changements du milieu***.
Des différenciations génétiques entre individus et entre populations peuvent ainsi se produire et conduire in fine à limiter la reproduction sexuée et les échanges de gènes entre individus/populations. Si ce type de différenciation génétique est suffisamment marquée, et donc que la reproduction sexuée n’est plus ou presque plus possible entre deux populations, on dit alors qu’il y a isolement reproducteur. A terme cela conduit à une spéciation (formation d’une nouvelle espèce).
Les processus évolutifs et de spéciation étant en cours tandis que les biologistes étudient les espèces, il est souvent compliqué de définir/délimiter une espèce. Néanmoins, les méthodes modernes de séquençage de l’ADN permettent de regrouper les individus qui possèdent des patrimoines génétiques proches. On contourne ainsi les difficultés liées aux critères morpho-anatomiques et d’interfécondité. On peut ainsi découvrir des groupes isolés génétiquement c’est à dire qui n’échangent plus de gènes et donc identifier de facto de nouvelles espèces.
* biologiste et ornithologue, à l’origine de la théorie synthétique de l’évolution (qui faisait la « synthèse » entre la génétique et la théorie de l’évolution proposée par Charles Darwin en 1859).
** on a longtemps cru que le mâle et la femelle Colvert étaient deux espèces différentes du fait de leur différences de plumage.
*** ces changements peuvent être biotiques (liées aux espèces présentes) ou abiotiques (paramètre physico-chimiques).
https://hal.archives-ouvertes.fr/ARINRA-COURENV/hal-01201814/file/C46Leguyader.pdf
Schéma-bilan du chapitre

Ce site a été réalisé par Mme et Mr ESTHER

Ce site a été réalisé par Mme et Mr ESTHER

Voici quelques exemples possibles de sujets de bac sur ce chapitre :
- aucun pour l’instant dans les annales (exercices de type 1 )
- aucun pour l’instant dans les annales (exercices de type 2)
- 1 seul exemple sur le suivi du groupe sanguin (Edu’modèles + Calc) exercices de type ECE1

rien pour l’instant !